By Victoria Osinski
What are Pattern recognition receptors?
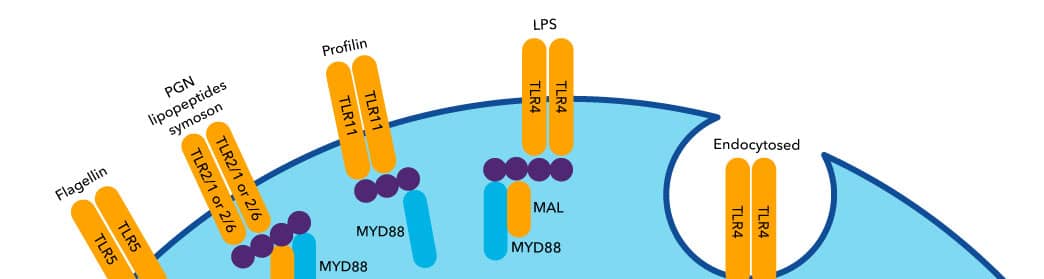
Pattern recognition receptors (PRRs) recognize and respond to Pattern- and Damage-associated molecular patterns (PAMP and DAMPs) in a number of different cell types. The PRR family can be broken down into sub-families of receptors (Table 1). While first identified in the context of infection, these receptors and signaling pathways are involved in many disease contexts including ischemia, cancer, atherosclerosis, and autoimmunity.2,16,18 Broadly, PRR signaling can regulate cell death1, proliferation, cell activation, and pro-inflammatory cytokine expression via nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) and interferon regulatory transcription factor (IRF) transcriptional regulation.16 While PRRs are typically thought of as receptors expressed on immune cells, they are also expressed on non-immune cells where they play important functional roles.
Table 1. Pattern recognition receptors1,2,7,10,11,14
| Receptor sub-family | Family members | Cellular localization |
|---|---|---|
| Toll-like Receptors (TLRs) | TLR1-13 | Plasma membrane and endosome (TLR3, TLR7, TLR8) |
| C-type lectin receptors (CLRs) | Mannose receptor 1, CLEC4E, Dectin-1, BDCA-2, DCAR, DCAR1, CLEC-1, CLEC9A, SIGNR3, MDL-1 | Plasma membrane |
| NOD-like receptors (NLRs) | 1. NLRAs: CIITA 2. NLRCs: NOD1/NOD2, NLRC3/NLRC4/NLRC5, NLRX1 3. NLRPs: NLRP1-12,14 4. NLRXs: NLRX1 5. NAIPs: NAIP |
Cytoplasm |
| RIG-I-like receptors (RLRs) | RIG-I, MDA5, DHX58 | Cytoplasm |
| Receptor for advanced glycation end-products (RAGE) | AGER (RAGE) | Plasma membrane, soluble |
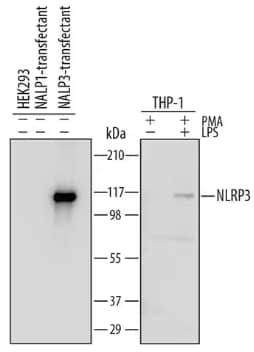
Western blot depicting lysates from HEK293 cell line either mock transfected, transfected with human NALP1 or transfected with NLRP3/NALP3 (left) and THP‑1 human acute monocytic leukemia cell line (right) untreated (-) or treated (+) with 20 ng/mL PMA for 72 hours and 1 µg/mL LPS for 24 hours. PVDF Membrane was probed with 0.5 µg/mL of Sheep Polyclonal Anti-NLRP3/NALP3 Antibody (AF6789) followed by HRP-conjugated Secondary Antibody (HAF016). Band was detected for NLRP3/NALP3 at approximately 115 kDa.
Innate immunity signaling in the vasculature
Multiple studies have shown that PRRs are expressed on endothelial cells8, which are cells that line the innermost layer of all blood vessels. Endothelial cells are important in the translocation of immune cells from the circulation to sites of infection or injury and are essential for vascular function and homeostasis. PRR signaling in endothelial cells influences multiple functions including their activation and endothelial cell barrier function.3-6,9,15 Many different PAMPs and DAMPs have been shown to bind to PRRs on endothelial cells including High mobility group box 1 (HMGB1), oxidized lipids, lipopolysaccharide (LPS), and Heat shock proteins (HSPs)3-6,9,15,16.
Examples of PRR activity in endothelial cells:
- Activation of TLR2 and TLR6 on endothelial cells leads to production of pro-inflammatory and pro-angiogenic molecules IL-8 and TNF-alpha via MyD88 and NFκB. Generally, TLR2 and TLR6 promote angiogenesis.
- TLR4 signaling upregulates E-selectin, PECAM-1, MCP-1, and IL-6 expression in endothelial cells3,5.
- Binding of DAMPs such as HMGB1 to RAGE in endothelial cells promotes maintenance of a tight endothelial barrier in blood vessels, which can be inhibited by increased presence of soluble RAGE. This mechanism is mediated by downstream beta-catenin and VE-cadherin signaling9.
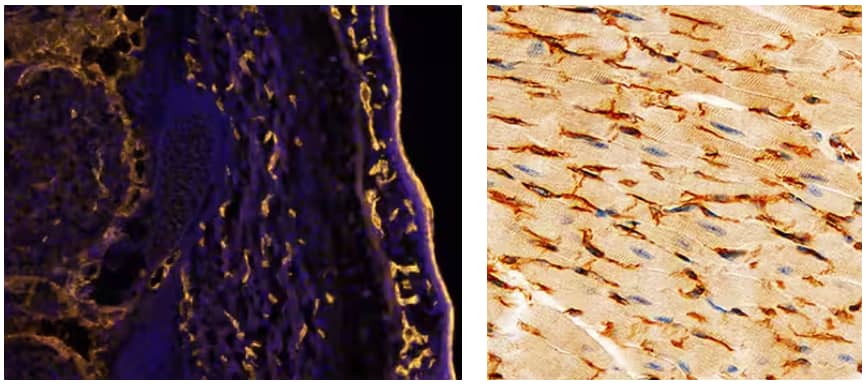
(Left) Fluorescent IHC staining of immersion fixed frozen sections of mouse embryo (E13.5) with Goat Polyclonal CD31/PECAM‑1 Antibody (AF3628) at 10 µg/mL overnight at 4 °C. Tissue was stained using the NorthernLights™ 557-conjugated Secondary Antibody (NL001) (yellow) and counterstained with DAPI (blue). Specific staining was localized to developing endothelium. (Right) IHC staining of immersion fixed paraffin-embedded sections of mouse heart with Goat Polyclonal VE‑Cadherin Antibody (AF1002) at 3 µg/mL for 1 hour at room temperature followed by incubation with the Anti-Goat IgG HRP Polymer Secondary Antibody (VC004). Tissue was then stained with DAB (brown) and counterstained using hematoxylin (blue). Specific staining was localized to cell membranes.
Beyond endothelial cells, there are many other non-immune cell types such as vascular smooth muscle cells and adipocytes whose functions are affected by PRR signaling. Innate immune signaling in the vasculature is an emerging and exciting field relevant to many different disease settings including cancer, infection, and cardiovascular disease.

Victoria Osinski, Doctoral Candidate
University of Virginia
Victoria studies cellular mechanisms regulating vascular growth during peripheral artery disease and obesity.
-
Amarante-Mendes, G. P. et al. (2018) Pattern Recognition Receptors and the Host Cell Death Molecular Machinery Frontiers in immunology
-
Cao X. (2016) Self-regulation and cross-regulation of pattern-recognition receptor signalling in health and disease Nature reviews. Immunology
-
Chen, K. et al. (2017) Ginkgolide B Suppresses TLR4-Mediated Inflammatory Response by Inhibiting the Phosphorylation of JAK2/STAT3 and p38 MAPK in High Glucose-Treated HUVECs Oxidative medicine and cellular longevity
-
Grote, K. et al. (2010) Toll-like receptor 2/6 stimulation promotes angiogenesis via GM-CSF as a potential strategy for immune defense and tissue regeneration Blood
-
Hijiya, N. et al. (2002) Possible involvement of toll-like receptor 4 in endothelial cell activation of larger vessels in response to lipopolysaccharide Pathobiology : journal of immunopathology, molecular and cellular biology
-
Into, T. et al. (2007) Pathogen recognition by Toll-like receptor 2 activates Weibel-Palade body exocytosis in human aortic endothelial cells The Journal of biological chemistry
-
Kato, H., & Fujita, T. (2015) RIG-I-like receptors and autoimmune diseases Current opinion in immunology
-
Khakpour, S. et al. (2015) Vascular endothelial cell Toll-like receptor pathways in sepsis Innate immunity
-
Jeong, J. et al. (2019) Soluble RAGE attenuates AngII-induced endothelial hyperpermeability by disrupting HMGB1-mediated crosstalk between AT1R and RAGE Experimental & molecular medicine
-
Motta, V. et al. (2015) NOD-like receptors: versatile cytosolic sentinels Physiological reviews
-
Saxena, S., & Jha, S. (2017) Role of NOD- like Receptors in Glioma Angiogenesis: Insights into future therapeutic interventions Cytokine & growth factor reviews
-
Sturtzel C. (2017) Endothelial Cells Advances in experimental medicine and biology
-
Tartey, S., & Takeuchi, O. (2017) Pathogen recognition and Toll-like receptor targeted therapeutics in innate immune cells International reviews of immunology
-
Tatematsu, M. et al. (2018) Extracellular RNA Sensing by Pattern Recognition Receptors Journal of innate immunity
-
Wang, L. et al. (2017) RAGE Plays a Role in LPS-Induced NF-κB Activation and Endothelial Hyperpermeability Sensors (Basel, Switzerland)
-
Wang, X., & Yi, F. (2015) Implication of pattern-recognition receptors in cardiovascular diseases Antioxidants & redox signaling
-
Wilhelm, I. et al. (2017) Role of pattern recognition receptors of the neurovascular unit in inflamm-aging American journal of physiology. Heart and circulatory physiology
-
Xu, J., & Sachdev, U. (2016) The Toll of Vascular Insufficiency: Implications for the Management of Peripheral Arterial Disease Journal of immunology research